





Cellular membranes are subject to constant dynamic modifications. Endocytosis and exocytosis are important processes by which membrane lipids and integral membrane proteins can be removed or inserted into the plasma membrane. PIs mark membrane areas for the recruitment of various proteins and in this fashion are involved in the control of endocytosis and exocytosis. Many proteins influencing endocytosis or exocytosis are known. Here we are investigating, which of these factors might be critically regulated by PIs. Through their polarized distribution PIs can, e.g., define membrane regions from which certain proteins are recycled or into which they are inserted. The control of the balance of recycling vs. re-insertion of PIN-proteins is an important example. Our experiments are designed to understand the influence of regulatory membrane lipids on abundance and polarization of plant integral membrane proteins at the molecular level.
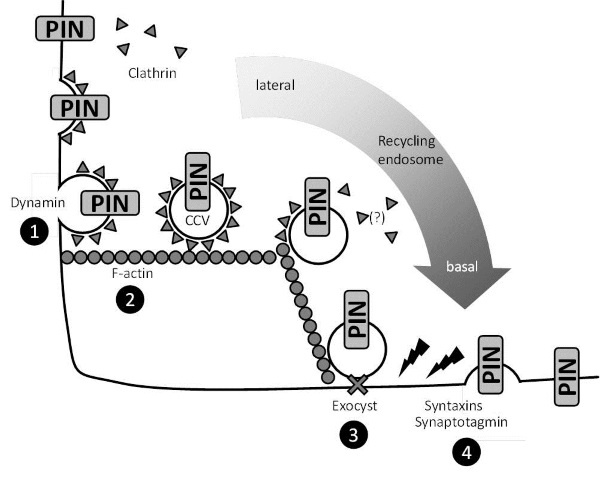
Endocytosis, exocytosis and the polarization of plant cells. Processes underlying, e.g., the polarization of PIN-proteins in cells of the Arabidopsis root tip: PIN proteins inserted in the plasma membrane (top left) are recycled via clathrin-mediated endocytosis (CME). Clathrin is initially recruited (“initiation”) to sites containing cargo proteins (“assembly”) to form clathrin-coated pits. Clathrin-coated vesicles (CCVs) containing PINcargo are cleaved off the plasma membrane (“scission”) by action of dynamins (1) and subsequently transported along F-actin-strands (2) towards the recycling endosome. The PIN-cargo is reinserted at other plasma membrane areas by first tethering the secretory vesicles to the plasma membrane via the exocyst complex (3) and fusing the vesicles into the target area of the plasma membrane by action of syntaxins and synaptotagmin (4). The role of PIs in these processes in plants is currently unclear.